ДомойДетские рецептыПримеры проявления закона взаимной индукции. Движение нервных процессов в коре больших полушарий
Примеры проявления закона взаимной индукции. Движение нервных процессов в коре больших полушарий
Деятельность коры головного мозга подчинена ряду принципов и законов. Основные из них впервые были установлены И.П.Павловым. В настоящее время некоторые положения павловского учения уточнены, развиты, а отдельные из них пересмотрены. Однако для овладения основами современной нейрофизиологии необходимо ознакомиться с фундаментальными положениями павловского учения.
Аналитико-синтетический принцип высшей нервной деятельности.
Как установлено И.П.Павловым, основным фундаментальным принципом работы коры больших полушарий головного мозга является аналитико-синтетический принцип. Ориентация в окружающей среде связана с вычленением отдельных ее свойств, сторон, признаков (анализ) и объединением, связью этих признаков с тем, что является полезным или вредным для организма (синтез). Синтез -это замыкание связей, а анализ -это все более тонкое отчленение одного раздражителя от другого.
Аналитико-синтетическая деятельность коры головного мозга осуществляется взаимодействием двух нервных процессов: возбуждения и торможения. Эти процессы подчинены следующим законам.
Закон иррадиации возбуждения. Очень сильные (так же, как и очень слабые) раздражители при длительном воздействии на организм вызывают иррадиацию -распространение возбуждения по значительной части коры больших полушарий.
Только оптимальные раздражители средней силы вызывают строго локализированные очаги возбуждения, что и является важнейшим условием успешной деятельности.
Закон концентрации возбуждения. Возбуждение, распространившееся из определенного пункта по другим зонам коры, с течением времени сосредоточивается в месте своего первичного возникновения.
Этот закон лежит в основе главного условия нашей деятельности -внимания (сосредоточенности сознания на определенных объектах деятельности).
При концентрации возбуждения в определенных участках коры мозга происходит его функциональное взаимодействие с торможением, это и обеспечивает нормальную аналитико-синтетическую деятельность.
Закон взаимной индукции нервных процессов. На периферии очага одного нервного процесса всегда возникает процесс с обратным знаком.
Если в одном участке коры сконцентрирован процесс возбуждения, то вокруг него индуктивно возникает процесс торможения. Чем интенсивнее сконцентрированное возбуждение тем интенсивнее и шире распространен процесс торможения.
Наряду с одновременной индукцией существует последовательная индукция нервных процессов -последовательная смена нервных процессов в одних и тех же участках мозга.
Только нормальное соотношение процессов возбуждения и торможения обеспечивает поведение, адекватное (соответствующее) окружающей среде.
Нарушение баланса между этими процессами, преобладание одного из них вызывает значительные нарушения в психической регуляции проведения.
Так, преобладание торможения, недостаточное взаимодействие его с возбуждением приводит к снижению активности организма. Преобладание возбуждения может выразиться в беспорядочной хаотической деятельности, излишней суетливости, снижающей результативность деятельности. Процесс торможения -это активный нервный процесс. Он ограничивает и направляет в определенное русло процесс возбуждения, содействует сосредоточению, концентрации возбуждения.
Торможение бывает внешним и внутренним. Так, если на животное внезапно подействует какой-либо новый сильный раздражитель, то прежняя деятельность животного в данный момент затормозится. Это внешнее (безусловное) торможение. В данном случае возникновение очага возбуждения по закону отрицательной индукции вызывает торможение других участков коры.
Одним из видов внутреннего или условного торможения является угасание условного рефлекса, если он не подкрепляется безусловным раздражителем (угасательное торможение). Этот вид торможения вызывает прекращение ранее выработанных реакций, если они в новых условиях становятся бесполезными.
Торможение возникает и при чрезмерном перевозбуждении мозга. Оно защищает нервные клетки от истощения. Этот вид торможения называется охранительным торможением.
На внутреннем виде торможения основана и аналитическая деятельность коры мозга, способность различать близкие по своим свойствам предметы и явления. Так, например, при выработке у животного условного рефлекса на эллипс оно вначале реагирует и на эллипс и на круг. Происходит генерализация, первичное обобщение сходных раздражителей. Но, если постоянно сопровождать предъявление эллипса пищевым раздражителем и не подкреплять предъявление круга, то животное постепенно начинает отчленять (дифференцировать) эллипс от круга (реакция на круг затормаживается). Этот вид торможения, лежащий в основе анализа, дифференцирования, называется дифференцировочным торможением. Оно уточняет действия животного, делает его более приспособленным к окружающей среде.
Р и с. 7. А-Иррадиация нервных процессов; Б - концентрация нервных процессов.
Без движения и взаимодействия основных нервных процессов -возбуждения и торможения
- невозможна высшая нервная деятельность. Движение нервных процессов - явление закономерное. И. П. Павлов открыл два основных закона движения нервных процессов в коре: закон иррадиации и концентрации и закон взаимной индукции.
Движение нервных процессов - явление закономерное. И. П. Павлов открыл два основных закона движения нервных процессов в коре: закон иррадиации и концентрации и закон взаимной индукции
Возбуждение или торможение коры больших полушарий
Возбуждение (или торможение)
, возникнув в каком-либо пункте коры больших полушарий, не остается в нем, а вначале иррадирует, т. е. распространяется на ближайшие нервные клетки, иногда захватывает обширные области коры (рис. 7, А). Спустя некоторое время наблюдается противоположное явление концентрации, т. е. сосредоточения нервного процесса в том месте, где он возник (рис. 7, Б). Поскольку клетки коры могут находиться в различном функциональном состоянии, иррадиация нервного процесса может встретить сопротивление со стороны противоположного нервного процесса, иррадирующего из другого пункта коры. Встреча противоположных процессов
вызывает их борьбу. Волна иррадирующего возбуждения «отгоняет» тормозной процесс из ближайших клеток в отдаленные пункты коры, но, если торможение становится достаточно сильным, что бывает при неподкреплении условного раздражителя, оно, распространяясь, в свою очередь «сгоняет» возбуждение к месту его возникновения. Очень убедительно явление иррадиации и концентрации нервных процессов доказывают широко известные эксперименты в лабораториях И. П. Павлова с раздражением кожного анализатора собаки посредством касалок.
Движение нервных процессов в коре больших полушарий имеет ряд закономерностей.
Возбуждение распространяется и концентрируется значительно быстрее, чем торможение. Скорость его движения измеряется секундами и долями секунд. Скорость движения процесса торможения
измеряется минутами, причем концентрация торможения происходит в 4-5 раз медленнее, чем иррадиация. Далее установлено, что движение нервных процессов
в коре
зависит от силы вызвавших их раздражителей, от функционального состояния коры мозга к моменту опыта и от уравновешенности возбуждения и торможения, что, в свою очередь, зависит от возраста и индивидуально-типологических особенностей организма.
Иррадиация возбуждения
Явление генерализации условий связи, о котором речь шла выше, объясняется иррадиацией возбуждения
по корковой части анализатора, а иногда и по близлежащим клеткам других анализаторов. Поэтому и возникает не специфический, обобщенный ответ организма на сходные раздражители. Генерализация возбуждения, по И. П. Павлову, имеет положительное и отрицательное значения. С одной стороны, это явление биологически оправдано. Агенты, на которые образуются натуральные условные рефлексы у животных, постоянно колеблются. Так, голос хищника, который служит сигналом опасности для животного-жертвы, колеблется в высоте, силе, составе в зависимости от напряжения голосового аппарата, расстояния, резонанса. Запах растения, который служит травоядному животному сигналом пищевого условного рефлекса, меняется в зависимости от влажности воздуха, расстояния, близости других запахов и прочих условий. Без генерализации животное не смогло бы все изменения раздражителя отнести к одному и тому же агенту и действовать соответственно его роли.
Отрицательное значение генерализации состоит в том, что иногда при широкой иррадиации возбуждения по клеткам коры в сферу обобщения включаются агенты, отдаленно сходные с основным сигналом; и это ведет к грубому неразличению, нежелательной путанице действий.
Явление генерализации условной связи есть явление простейшего коркового синтеза.
За генерализацией условных рефлексов следует их специализация
Т. е. отчетливое выделение сигнального раздражителя из массы сходных с ним агентов. Она объясняется концентрацией нервных процессов
в определенных пунктах коры, что вызвано дифференцировочным торможением. Явление специализации условной связи есть явление коркового анализа. Специализированные условные рефлексы могут вступать в связь друг с другом, образуя сложные функциональные системы. Такой вторичный синтез по своему уровню выше, чем первичная генерализация. Он основан на избирательной генерализации. Аналитико-синтетическая деятельность коры развивается у животного в процессе усложнения его связи с окружающим миром, у человека - в процессе обучения и воспитания.
Изучение закономерностей коркового торможения позволило раскрыть физиологию сна. Сон, по учению И. П. Павлова, имеет условно-рефлекторную природу и возникает в результате широкой иррадиации торможения, которая охватывает всю кору полушарий и спускается ниже- в подкорку и даже средний мозг. Сонное торможение может быть вызвано разными причинами: понижением уровня работоспособности корковых клеток в результате длительного и напряженного их функционирования, ограничением поступающих в кору раздражений извне (длительное пребывание в темноте, в тишине при неподвижности или ритмических движениях тела может вызвать сон даже в том случае, если человек не утомлен) и привычкой засыпать в определенное время. В экспериментальной обстановке сон может быть вызван длительным, непрерывным действием какого-либо раздражителя без подкрепления его безусловным. В этом случае угасательное торможение, иррадируя, переходит в сонное.
Сонное торможение распространяется по коре с неравномерной скоростью и силой.
Некоторые группы нервных клеток, в которых локализован стойкий процесс возбуждения
, могут остаться не заторможенными и во время сна. Образуются так называемые «сторожевые пункты», что ведет к немедленному просыпанию под действием определенных сигналов, даже слабых. Таким бывает сон кормящей матери, которая сразу же просыпается на слабые звуки, исходящие от ребенка (стон, затрудненное дыхание, легкое движение ребенка).
Причины сна могут быть и иными. Сон можно вызвать резким ограничением внешних раздражителей, а также электрическим раздражением специальных подкорковых центров. Сон необходим организму для восстановления работоспособности нервных клеток.
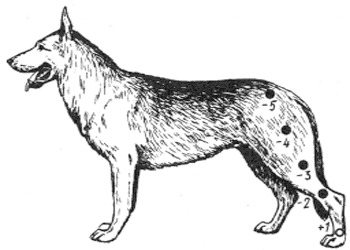
Р и с. 8. Одновременная индукция: А - положительная; Б - отрицательная
Возбуждение и торможение взаимно индуцируют
, т. е. вызывают и усиливают друг друга. Возбуждение вызывает торможение и наоборот. Чем сильнее возбуждение, тем сильнее будет и вызванное им торможение. Различают два вида индукции: положительную и отрицательную, каждая из которых может быть одновременной и последовательной. Если исходным процессом является возбуждение, которое по индукции вызывает торможение, это - – отрицательная индукция (рис. 8, Б), а если торможение вызывает возбуждение, это - положительная индукция (рис. 8, Л), при одновременной индукции нервные процессы располагаются в разных пунктах коры и существуют вместе, а при последовательной индукции (рис. 8, А, Б) нервные процессы
сменяют друг друга в одном и том же пункте коры.1 Приведем примеры различных видов индукционных отношений в коре.
При большой сосредоточенности слухового внимания человек сидит неподвижно, не замечает ничего, что не относится к объекту его внимания. Процесс возбуждения
сконцентрирован в определенных корковых клетках слухового анализатора, а вокруг них временно разлито торможение. Это одновременная отрицательная индукция. Но вот звуки, к которым прислушивался человек (например, речь учителя), прекратились. Теперь в рабочих клетках слухового анализатора возбуждение сменяется торможением. Это - последовательная отрицательная индукция. Если ученики на уроке физики самостоятельно решали задачи, а затем учитель предложил им наблюдать за демонстрацией физического опыта, то такая смена умственной деятельности влечет за собой временный отдых, торможение рабочих клеток определенных полей мозга после длительного их возбуждения. Это тоже последовательная отрицательная индукция
.
Примером одновременной положительной индукции может служить явление контраста при восприятии.
Так, светло-серый фон вокруг черного квадрата по контрасту кажется белым. От черного квадрата нет световых раздражений. В соответствующих корковых клетках зрительного анализатора возникает тормозной процесс, который по индукции усиливает процесс возбуждения, возникший в соседних клетках от восприятия светло-серого фона. Создается иллюзия более яркого освещения этого фона, чем оно есть в действительности. Второй пример. Однотонная тихая речь учителя на уроке, не сопровождаемая демонстрацией наглядных пособий или опытов и не содержащая ярких описаний, очень быстро утомляет школьников, особенно детей младшего возраста. Их внимание становится рассеянным. В утомленных нервных клетках
рече-слуховой области коры возникает процесс торможения, который по индукции усиливает возбуждение соседних нервных Клеток зрительного, слухового и двигательного анализаторов, вызываемое действием слабых раздражений: ребенок теперь замечает случайный скрип парты, шелест бумаги сзади, покашливание; разглядывает свои руки и предметы, лежащие на парте, сидящих впереди учеников; перебирает в карманах или в парте какие-либо знакомые вещи и т. д. Ориентировочные рефлексы на посторонние слабые раздражители усиливаются именно потому, что основной раздражитель - голос учителя - вызвал стойкое торможение в рече-слуховой области коры. Это - одновременная положительная индукция. В качестве примера последовательной положительной индукции можно привести тот же факт со скучным уроком: после долгого вынужденного сидения в классе даже дисциплинированные дети и подростки проводят довольно шумные перемены. Длительное торможение двигательных реакций
сменилось повышенной двигательной активностью. Индукционные отношения основных нервных процессов существуют также между корой и ближайшей подкоркой. При сильных эмоциях (гнев, страх, отчаяние) возбужденная подкорка вызывает по индукции торможение корковых нервных связей, прежде всего второсигнальных, Этим объясняется недостаточная разумность некоторых действий эмоционально возбужденного человека. Возможно и обратное отношение.
Задачи раздела:
дать
характеристику процессов иррадиации и
концентрации возбуждения и торможения,
рассмотреть закон взаимной индукции и его
проявление, изучить явление доминанты и ее роль в
психических процессах, познакомиться с
физиологическими основами и теориями сна и
сновидений, гигиеной сна.
Урок 1. ИРРАДИАЦИЯ И КОНЦЕНТРАЦИЯ
НЕРВНЫХ ПРОЦЕССОВ
Оборудование:
таблицы, схемы и
рисунки, иллюстрирующие процессы иррадиации и
концентрации возбуждения и торможения.
ХОД УРОКА
I. Изучение нового материала
Динамика нервных процессов в сети
нейронов
Вся сложная и разнообразная
деятельность высших отделов нервной системы
построена на работе двух основных нервных
процессов – возбуждения и торможения. Протекая в
подвижных пространственных и временных
отношениях друг с другом, эти процессы то
разливаются (иррадиируют), то сосредоточиваются
(концентрируются) в определенных пунктах коры, то
возбуждение порождает торможение (отрицательная
индукция), то торможение дает начало возбуждению
(положительная индукция).
Непрерывное взаимодействие
движущихся и вызывающих друг друга
возбудительных и тормозных процессов создает в
высших отделах мозга чрезвычайно тонкую мозаику,
колеблющийся узор из переплетения возбужденных
и заторможенных нейронов. Такие мозаики лежат в
основе как различных актов поведения, так и их
торможения в явлениях сна.
Иррадиация торможения
Возбуждение или торможение, возникшее
в какой-либо клетке или группе клеток мозга,
всегда склонны к распространению.
Распространение нервного процесса из очага его
возникновения на окружающие нервные клетки
называется иррадиацией
(от лат. irradiare
–
сиять).
Иррадиацию условного торможения
удобно наблюдать в кожном анализаторе.
Значительная площадь этого анализатора является
как бы увеличивающим зеркалом, в котором можно
ясно видеть, как по последовательно
расположенным проекционным полям будет
иррадиировать тормозное состояние, например
дифференцировочное торможение.
Рис. 1. Опыт с иррадиацией
дифференцировочного торможения по корковым
клеткам кожного анализатора:
0 – дифференцировочный раздражитель; 1, 2, 3, 4 –
положительные условные раздражители
(прикладываются к пунктам кожи ноги на
расстоянии от дифференцировочного раздражителя
соответственно на 3, 9, 15 и 22 см)
Иррадиацию дифференцировочного
торможения обнаружили в следующем опыте
(рис. 1). Вдоль задней ноги собаки от стопы до
бедра наклеивали пять «касалок» – приборов для
механического раздражения кожи. Четыре верхние
касалки использовали для выработки условных
пищевых слюноотделительных рефлексов и
добивались одинаковых слюноотделительных
эффектов от этих раздражителей. Нижняя касалка
служила дифференцировочным раздражителем и
применялась без подкрепления едой, пока не
переставала вызывать даже малейшее
слюноотделение. Если теперь, вслед за
применением дифференцировочной касалки,
испробовать положительные раздражители, то
оказывается, что слюногонное действие последних
претерпевает закономерные изменения.
Каждый раз, когда дифференцировочная
касалка создавала очаг торможения, начинали
изменяться и соседние положительные рефлексы.
Следовательно, торможение выходит за пределы
своего очага и захватывает соседние клетки
анализатора, в данном случае те, на которые
проецируются пункты положительных касалок.
В одинаковых условиях условные
рефлексы, связанные с положительными касалками,
изменяются по-разному. Так, рефлекс, связанный с
ближайшим пунктом (касалка 1), оказался полностью
заторможенным. Рефлекс, связанный с пунктом,
расположенным несколько дальше (касалка 2), был
только уменьшен. Рефлексы, связанные с пунктами,
расположенными еще дальше, не только не испытали
торможения, но даже усилились. Следовательно,
иррадиирущее торможение оказывает тем более
сильное воздействие на клетки анализатора, чем
ближе они находятся к тормозном очагу.
Каждый, кому приходилось играть в мяч,
знает, как легко обмануть партнера, сделав
несколько обманных движений мячом. После ряда
таких бросков партнер не только не пытается
поймать мяч, но даже не сходит с места, не меняет
позы. Торможение, наступившее вследствие
угасания условного рефлекса на бросок мяча,
распространилось на многочисленные нервные
центры. Этот пример также иллюстрирует
иррадиацию торможения.
Концентрация торможения
После широкой иррадиации наступает
сосредоточение, концентрация торможения
в
месте его возникновения. Этот процесс тоже
удобно проследить на примере
дифференцировочного торможения в кожном
анализаторе. Опыты проводили так же, как и при
наблюдении иррадиации, но положительные
рефлексы на раздражение каждого участка кожи
испытывали в различные сроки после окончания
действия тормозного раздражителя. При помощи
такого приема можно увидеть, как сначала далеко
распространившееся тормозное состояние
начинает сосредоточиваться, возвращаясь к
исходному пункту.
При концентрировании торможение
проходит в обратной последовательности все те
пункты проекционных полей анализаторов, которые
оно захватывало в своем поступательном движении.
Что представляет собой процесс
торможения? Возможны два варианта. При первом
распространившееся торможение рассеивается,
затухает на периферии и занимаемая им территория
постепенно уменьшается. При втором происходит
подъем обратной волны торможения к тому месту,
откуда оно распространилось. Последнее более
вероятно, так как, например, упрочение
дифференцировки сопровождается усилением
тормозного процесса.
Следовательно, концентрация
торможения связана не с рассеиванием и
ослаблением, а с его сосредоточением и усилением.
Скорость иррадиации и концентрации
торможения
На основании ряда опытов с кожным
анализатором удалось измерить скорость
иррадиирования тормозного состояния. Оказалось,
что процесс иррадиирования торможения по
нервным клеткам коры протекает очень медленно.
Для прохождения области одного только кожного
анализатора торможению требуются минуты.
Абсолютные величины времени
концентрации тормозного процесса, как и времени
его иррадиации, сильно зависят от индивидуальных
особенностей подопытных животных, однако их
соотношение оказалось довольно постоянным у
всех испытуемых собак. Как правило,
иррадиирование происходит в 4–5 раз быстрее, чем
последующее концентрирование.
Иррадиация и концентрация
возбуждения
Опыт, показывающий иррадиацию
возбудительного процесса, в некоторых
отношениях напоминает описанные опыты с
иррадиацией торможения.
У собаки вдоль задней ноги от плюсны до
таза на примерно одинаковом расстоянии друг от
друга приклеивали пять касалок. На действие
самой нижней касалки (касалка 1) вырабатывали
условный рефлекс слюноотделения, подкрепляемый
вливанием в рот собаке подкисленной воды. При
первом испытании и другие сходные раздражители
(касалки 2, 3, 4 и 5) вызывали слюноотделение. Для
выработки дифференцированных реакций от касалок
применли многократно касалку 1 с
подкреплением, а остальные касалки – без
подкрепления. Теперь только касалка 1 вызывала
слюноотделение, а остальные превратились в
тормозные сигналы.
После такой подготовки приступали к
главной части опыта. Включали положительную
касалку 1 на 15 с и сразу после ее выключения
действовали касалкой 2. Однако ее действие тоже
вызывало слюноотделение. Это означало, что пункт
кожного анализатора, под касалкой 2, обычно
находящийся в тормозном состоянии, сразу после
возникновения очага возбуждения в пункте, под
касалкой 1, тоже оказывался в возбужденном
состоянии. Иначе говоря, возбуждение из пункта
под касалкой 1 в это время распространилось на
пункт под касалкой 2. Если так же испытать и
какой-либо другой, более удаленный, пункт кожного
анализатора, то можно судить о районе такой
иррадиации. Таким образом, иррадиирущее
возбуждение по мере удаления от очага своего
развития постепенно ослабевает (рис. 2).
Рис. 2. Опыт с иррадиацией возбуждения
по корковым клеткам кожного анализатора:
1 – положительный условный раздражитель; 2, 3, 4, 5 –
дифференцировочные раздражители
Опыты показали, что иррадиация
возбуждения в коре мозга происходит гораздо
быстрее, чем иррадиация торможения, и требует
менее 1 с для распространения по области кожного
анализатора.
Через некоторое время после
положительного сигнала соседние пункты
анализатора вновь оказываются в прежнем
тормозном состоянии. Это означает, что волна
возбуждения уже успела разлиться по коре и вновь
сосредоточиться в исходном пункте.
Сходные картины можно наблюдать и в
жизни человека. Ребенку прижгли рану на руке
йодом. Сначала он отдернул руку, затем стал ею
махать, потом подпрыгивать, плакать, кричать.
Возбуждение, возникшее в одном пункте коры,
распространилось на другие. Оно охватило
обширные участки коры, подкорковые центры.
В процессе обучения какому-либо навыку
человек сначала совершает большое количество
ненужных движений, и только после более или менее
длительной практики его движения становятся
экономичными, координированными. Иррадиация
возбуждения уступает место концентрации,
вследствие которой возбуждение оказывается
стянутым в определенные участки.
Благодаря иррадиации возбуждения
животное может реагировать не только на тот
условный раздражитель, на который вырабатывался
условный рефлекс, но и на сходные раздражители.
Кошка обнаружила мышь по писку и поймала ее. Писк
мыши сделался условным раздражителем. Но только
ли на этот звук будет реагировать кошка?
Оказывается, нет. Благодаря иррадиации
возбуждения она будет реагировать на массу
сходных звуков: писк птенцов, стрекотание
кузнечика и т.д. Вполне возможно, что некоторые из
них окажутся небесполезными. Иррадиация делает
условный рефлекс обобщенным, или, как говорят,
генерализованным. Только спустя некоторое время
после образования этого рефлекса благодаря
дифференцировочному торможению животное
научится отличать истинные сигналы от ложных.
Благодаря концентрации возбуждения уловный
рефлекс становится специализированным.
Таким образом, способностью к
иррадиации и концентрации обладают и процесс
возбуждения, и процесс торможения.
II. Закрепление знаний
Обобщающая беседа по ходу изучения
нового материала.
III. Домашнее задание
Изучить параграф учебника (понятия
иррадиации и концентрации нервных процессов,
иррадиация и концентрация торможения и их
скорость, иррадиация и концентрация
возбуждения).
Урок 2–3. ИНДУКЦИЯ НЕРВНЫЙ ПРОЦЕССОВ
Оборудование
: таблицы, схемы и
рисунки, иллюстрирующие процессы иррадиации и
концентрации возбуждения и торможения, а также
процессы положительной и отрицательной
индукции, явление доминанты.
ХОД УРОКА
I. Проверка знаний
Работа по карточкам
Докажите, что на ранних стадиях
выработки условного рефлекса происходит
иррадиация возбуждения в коре полушарий
большого мозга.
1. Общая характеристика процессов
иррадиации и концентрации возбуждения и
торможения.
2. Характеристика иррадиации торможения.
3. Характеристика концентрации торможения.
4. Характеристика иррадиации и концентрации
возбуждения.
5. Скорость иррадиации и концентрации тормозных и
возбудительных процессов.
II. Изучение нового материала
Положительная индукция нервных
процессов
Движение основных процессов ВНД
определяется не только свойствами иррадиации и
концентрации, но и свойствами их взаимной
индукции. Индукцией
(от лат. induction
–
возбуждение) называется свойство каждого из
основных нервных процессов вызывать вокруг себя
и после себя противоположный процесс.
Явление, при котором процесс
торможения порождает процесс возбуждения,
называется положительной индукцией
.
Явление положительной индукции было
выявлено в специальных опытах на примере,
связанном с дифференцировочным торможением. Так,
у собаки вырабатывали условный пищевой рефлекс
слюноотделения, в котором сигналом служило
раздражение касалкой кожи передней лапы. Другая
касалка была установлена на задней ноге. Ее
применяли без подкреплений, так что скоро она
действовала как тормозной дифференцировочный
раздражитель. На включение дифференцировочной
касалки слюноотделение не наступало, однако
испробованный сразу после нее положительный
раздражитель давал резко усиленный рефлекс.
Измерение силы условного рефлекса по
количеству слюны обнаруживает, что торможение в
пункте задней лапы усилило условное возбуждение
в пункте передней лапы почти на 50%. Следовательно,
в данном случае произошла положительная
индукция из очага торможения в очаг возбуждения.
С положительной индукцией мы
встречаемся довольно часто в жизни. У малыша,
утомившегося в течение дня, в коре больших
полушарий начинают развиваться процессы
торможения, поскольку данный отдел обладает
наименьшей выносливостью. Торможение в коре по
закону положительной индукции вызывает
возбуждение подкорковых центров, в частности
тех, с которыми связаны эмоции. Ребенок начинает
либо веселиться, либо капризничать. Часто
положительные и отрицательные эмоции сменяют
друг друга: ребенок то плачет, то вновь начинает
смеяться.
Примерно то же бывает у опьяневшего
человека. Алкоголь вызывает наркотическое
торможение в коре, что приводит к возбуждению
подкорковых центров вследствие положительной
индукции. Эмоциональные реакции усиливаются,
человек переходит в состояние болезненной
веселости – эйфории, которая нередко сменяется
сильной тоской. Поведение становится
ненормальным, нередко агрессивным. Теряется
критическое отношение к обстановке, опьяневший
человек не может оценить степень риска. Ему все
кажется доступным и возможным. Это делает
пьяного человека социально опасным.
Отрицательная индукция нервных
процессов
Процесс, в ходе которого возбуждение
вызывает торможение, называется отрицательной
индукцией.
Явление отрицательной индукции можно
продемонстрировать в следующем опыте. У собаки
образован условный пищевой рефлекс на метроном с
частотой 120 ударов в минуту. К этому
положительному раздражителю выработана
дифференцировка метронома с частотой 60 ударов в
минуту. Как известно, дифференцировку очень
легко разрушить, если начать сопровождать
дифференцировочный раздражитель подкреплением.
И действительно, после того, как несколько раз
метроном с частотой 60 ударов в минуту применили с
подкреплением, он сам начал вызывать
слюноотделение. Это простой и безотказный способ
уничтожения тормозного очага.
После разрушения дифференцировки
применяют с подкреплением один метроном с
частотой 120 ударов в минуту. В результате
используемый вслед за ним метроном с частотой 60
ударов в минуту, который только что вызывал
слюноотделение, сразу теряет свое действие.
Дифференцировка при этом восстанавливается, что
связано с возникновением очага возбуждения. Этот
очаг отрицательно индуцировал, т.е. затормозил
клетки пункта метронома с частотой 60 ударов в
минуту, и индуцированное торможение усилило
остатки дифференцировочного.
Приведем пример отрицательной
индукции из жизни человека. Ребенку дали суп, он с
аппетитом начал его есть, но тут включили
телевизор, и ребенок застыл с поднятой ложкой.
Произошло знакомое внешнее торможение: сильное
возбуждение зрительных центров затормозило
пищевой центр.
Доминанта и ее роль в психических
процессах
Поведение во многом определяется
потребностями. В том случае, когда одна из
потребностей перерастает в сильное желание, она
может подчинить себе все остальное. Известный
физиолог А.А. Ухтомский обнаружил, что в
нервной системе, в частности в головном мозге,
могут возникать сильные очаги временного
возбуждения. Эти временно господствующие очаги
возбуждения в ЦНС, обладающие повышенной
возбудимостью ко всем приходящим в них
раздражениям и способные оказывать тормозящее
влияние на деятельность других нервных центров,
были названы доминантами
(от лат. dominantis
–
господствующий).
В условиях доминанты легко образуются
условно-рефлекторные связи между сигнальным
раздражителем и безусловным подкреплением.
Доминанты способны не только оказывать
интенсивную отрицательную индукцию на соседние
участки, вследствие чего достигается
значительная заторможенность тех полей, которые
не относятся к доминанте, но и возбуждения,
вызванные раздражителями, не относящимися к
доминанте, меняют свое обычное направление.
Нервные импульсы, вместо того, чтобы двигаться по
своему традиционному пути, идут в сторону
доминантного очага. Доминанта как бы притягивает
их и усиливается за их счет.
Например, если после выработки у
морской свинки условного жевательного рефлекса
на постукивание по столу, вместо постукивания
сказать любую фразу, животное начнет жевать.
Морская свинка начнет жевать, когда услышит
голос, и прекратит жевание, когда вы кончите
говорить. Любое раздражение – слуховое,
тактильное, зрительное – будет вызывать у нее
жевательные движения без предварительной
выработки. При выработке пищевого условного
рефлекса у морской свинки была создана
доминанта. Новые раздражители (голос человека и
др.) теперь без всякой выработки оказываются
связанными с пищевым возбуждением. Это
происходит потому, что нервные импульсы,
появившиеся под влиянием этих раздражителей,
меняют свой обычный путь, иррадиируя в сторону
доминантного очага возбуждения, как бы
притягиваются им. Они усиливают доминантное
возбуждение, что мы и видим по появлению
жевательной реакции.
А.А. Ухтомский полагал, что
доминировать могут целые системы рефлексов.
Доминанта лежит в основе таких психических
процессов, как внимание, сосредоточенность,
способность к волевым усилиям. Благодаря
доминанте человек целиком «уходит» в работу,
ничто его не отвлекает, он не слышит, когда к нему
обращаются. Внимание концентрируется на том, что
он делает. Алкоголик в состоянии запоя не может
думать ни о чем, кроме выпивки. Нередко он не в
силах контролировать свои поступки и становится
опасным для окружающих.
Однако в некоторых случаях появление
длительно не затухающих очагов доминантного
возбуждения может стать причиной различных
душевных заболеваний. Подобного рода застойные
очаги патологического возбуждения наблюдал
И.П. Павлов. Они являются одной из причин, в
силу которых душевнобольные люди неправильно
оценивают события и ненормально на них
реагируют.
Функциональная мозаика в высших
отделах нервной системы
Взаимодействие иррадиирущих и
индуцированных нервных процессов создает
необычайно сложное и меняющееся от момента к
моменту их уравновешивание и территориальное
разграничение. В результате возбуждение и
торможение образуют дробный рисунок подвижной
мозаики, непрерывно меняющей свои очертания
(рис. 3).
Рис. 3. Перераспределение очагов
активности в коре мозга кролика при выработке
длительного условного рефлекса на зрительное
раздражение
В свое время И.П. Павлов говорил о
том, какую замечательную картину вспыхивающих и
затухающих, непрерывно перемежающихся мерцаний
мы увидели бы на поверхности мозга, если бы его
возбужденные пункты светились. Это стало
возможным при изучении движения нервных
процессов по коре полушарий большого мозга с
помощью методики электроэнцефалоскопии
.
Электроэнцефалоскоп позволяет наблюдать
мозаику электрической активности коры мозга при
одновременном отведении из 100 ее пунктов и
воспроизводит на экране телевизора непрерывно
возникающие и меняющиеся подвижные картины,
которые фиксируются киносъемкой. Такой
«телевизор» мозга значительно расширяет
возможности объективного изучения
пространственной динамики активности коры при
условно-рефлекторной деятельности.
III. Закрепление знаний
Лабораторная работа № 4. «Изучение
явления взаимной индукции процессов возбуждения
и торможения»
Оборудование:
рисунки
двойственных изображений.
ХОД РАБОТЫ
1. Рассмотрите рисунок «ваза – два
профиля» (рис. 4). Найдите на нем два черных
профиля, обращенных лицом друг к другу, и белую
вазу (она находится между профилями).
2. Почему, когда видна ваза, исчезают
профили, а когда мы видим профили, пропадает
изображение вазы? (Причина заключается в том,
что один из конкурирующих образов затормаживает
появление второго, т.е. имеет место отрицательная
индукция: возбуждение наводит торможение).
3. Смотрите на рисунок «ваза – два
профиля» до тех пор, пока изображения не начнут
сменять друг друга: будет видна то ваза, то два
профиля. Объясните данное явление. (Когда мы
видим вазу, то возбужден комплекс нервных связей,
воспринимающих ее, а комплекс связей,
воспринимающих два профиля заторможен. Однако по
закону последовательной индукции после одного
процесса появляется противоположный, и
возбуждение сменяется торможением в одном
комплексе нервных связей, и торможение сменяется
возбуждением в другом
).
5. Вывод: с каким законом вы столкнулись
при выполнении лабораторной работы?
IV. Домашнее задание
Изучить параграф учебника
(положительная и отрицательная индукция, явление
доминанты, функциональная мозаика в сети
нейронов).
Урок 4–5. СОН ЧЕЛОВЕКА И ЕГО
ХАРАКТЕРИСТИКА. ТЕОРИИ СНА. СНОВИДЕНИЯ
Оборудование
: таблицы, схемы и
рисунки, иллюстрирующие процессы положительной
и отрицательной индукции, явление доминанты,
стадии сна.
ХОД УРОКА
I. Проверка знаний
Работа по карточкам
1. Приведите примеры проявления
закона взаимной индукции возбуждения и
торможения.
2. Каково значение явления доминанты в жизни
человека?
Устная проверка знаний по вопросам
1. Закон взаимной индукции нервных
процессов. Положительная индукция.
2. Отрицательная индукция.
3. Явление доминанты.
4. Функциональная мозаика в высших отделах
нервной системы.
II. Изучение нового материала
Сон человека и его физиологическое
значение
Явления природы нередко строго
периодичны: сменяются времена года, фазы Луны,
день и ночь. Живые организмы приспособились к
этим изменениям. Активное поведение людей
преимущественно приурочено к дневным часам.
Ночью наступает сон, и уставшие люди за ночь
отдыхают.
Сон –
периодически наступающее
физиологическое состояние у позвоночных
животных и человека, характеризующееся почти
полным отсутствием реакций на внешние
раздражения, уменьшением активности ряда
физиологических процессов.
Примерно треть своей жизни человек
проводит во сне. Чередование сна и бодрствования
– необходимое условие жизнедеятельности
человеческого организма. Без сна жизнь
невозможна. Так, в опыте собаки жили без пищи 20–25
суток и теряли 50% массы, а без сна 10–12 суток, хотя
их масса убывала всего на 5–13%.
Сколько же времени нужно спать? Это
зависит от возраста. Новорожденный спит почти
все время, он бодрствует только 2–3 ч в сутки;
шестимесячный малыш спит около 14 ч, годовалый –
13 ч. В четырехлетнем возрасте дети спят до
12 ч в сутки, в семилетнем – 11 ч, в
десятилетнем – 10 ч. Пятнадцатилетние
подростки должны спать 9 ч в сутки, а начиная с
17–18 лет продолжительность сна может
равняться в среднем 7–8 ч. В старческом
возрасте обычно спят меньше. Вместе с тем
длительность сна может быть различна у разных
людей. Из жизнеописания Петра I следует, что он
спал не более 5–6 ч, и этого было ему
достаточно. Описаны также многочисленные случаи,
когда человек довольствовался еще более
ограниченным временем сна.
Постоянное недосыпание может вызвать
головные боли, повышенную утомляемость и
способствовать ухудшению памяти, появлению
нервных и других болезней. Длительный сон так же
вреден, как и длительное бодрствование. Сном
нельзя запастись впрок.
Мозг поддерживается в бодрствующем
состоянии за счет импульсов, поступающих от
рецепторов тела. При прекращении или резком
ограничении их поступления в кору развивается
сон. Сон также развивается при действии на
корковые клетки длительной или чрезмерной силы
раздражителей. При этом в клетках коры
развивается торможение, имеющее охранительное
значение. Оно обеспечивает коре полушарий
большого мозга условия для восстановления
работоспособности во время сна.
В настоящее время установлено
существование в стволовой части головного мозга
образований, оказывающих влияние на наступление
бодрствования и сна. На бодрствование
существенное влияние оказывает ретикулярная
формация, а на сон – таламус.
По поводу физиологического значения
сна
существуют разные предположения, которые
условно можно свести в следующие группы.
Восстановление специфического
метаболизма нервных клеток мозга, которое
обеспечивает его полноценную деятельность в
состоянии бодрствования. И.П. Павлов полагал,
что наступающее в течение напряженной дневной
работы «истощение» корковых клеток вызывает сонное
торможение
, в течение которого
восстанавливается их работоспособность. По
Павлову, «сон – это общее торможение,
наступающее тогда, когда клетки мозга нуждаются
в отдыхе». Сон охраняет мозг от перенапряжения,
во сне перерабатывается информация, накопленная
за день, рождаются новые идеи.
Приспособление к неблагоприятным
условиям деятельности. Животные, ведущие дневной
образ жизни, становятся беспомощными ночью, так
как не ориентируются в темноте и могут стать
легкой добычей ночных хищников. В свою очередь
последние оказываются в аналогичном положении
днем. Сон обеспечивает не только отдых, но и
безопасность путем защитной неподвижности в
укромном месте. Это один из видов инстинктивного
приспособительного поведения.
Упорядочение процессов обработки и
хранения информации. Значение сна для состояния
памяти понимают двояко. Ряд ученых считают, что
происходит устранение накопившейся за день
«ненужной» информации, наступает
«дезинтеграция» памяти. Такую подготовку мозга к
восприятиям следующего дня сравнивают со
стиранием информации в ячейках памяти
компьютера. Другие, наоборот, считают, что во
время сна происходит консолидация памяти,
переход кратковременной в долговременную.
Высказывают также предположения о переработке
информации, которую мозг не успел переработать в
течение дня.
Восстановление согласованности
временного течения функций организма.
Бесчисленное множество биохимических реакций
выстраивается в сложную систему обеспечения
функций клеток, тканей и органов. Согласование во
времени этих взаимосвязанных, периодически
изменяющихся функций является необходимым
условием нормальной жизни организма.
Таким образом, сон – это защитное
приспособление организма, предупреждающее
переутомление нервной системы.
Характеристика стадий сна человека
По показателям электрической
активности мозга ночной сон можно разделить на два
периода (фазы)
:
медленноволновый
(медленный сон);
парадоксальный,
или
быстроволновый
(быстрый сон).
Время сна дифференцировано на
медленный и быстрый сон главным образом для
пластических восстановительных процессов,
обработки накопившейся информации и
консолидации долговременной памяти.
Во время сна меняется физиологическая
активность организма: расслабляется
мускулатура, снижаются кожная чувствительность,
зрение, слух, обоняние, затормаживаются условные
рефлексы. Дыхание во сне редкое, величина
кровяного давления, частота сердечных
сокращений снижены. Но сон – это не
бездеятельное состояние нервной системы. Во
время сна в нейронах происходят электрические
разряды, но характер электрической активности
меняется. Некоторые реакции у спящего человека
усиливаются: расширяются сосуды кожи, краснеет
лицо, повышается тонус некоторых мышц,
усиливае6тся секреция желудочных и кишечных
желез, более интенсивно идет всасывание,
активируются многие синтетические процессы.
Динамику электрической
активности мозга в процессе развития и течения
сна у человека изучали многие исследователи.
Была предложена классификация стадий сна на
основе изменений уровня сознания и формы
электроэнцефалограммы. К основным стадиям
развития естественного сна у человека относят
(рис.
6):
стадия А
– исходная для
засыпания. В нейронах головного мозга
преобладают электрические волны с частотой 8–12
колебаний в секунду, что характерно для
состояния спокойного бодрствования;
стадия В
– сонливость.
Преобладают низковольтные колебания разной
частоты;
стадия С
– поверхностный сон. В
электрической активности мозга появляются
веретенообразные группы колебаний с частотой
12–14 колебаний в секунду и отдельные медленные
волны;
стадия D
– углубление сна.
Появляются гигантские (200–300 мкВ) медленные волны
(1–3 колебаний в секунду);
стадия Е
– глубокий сон,
непрерывные ряды медленных волн. Медленный сон
сопровождается урежением дыхания, пульса,
расслаблением мышц. Он характеризуется
сновидениями и грезами;
стадия Р (парадоксальная)
–
глубокий сон, сопровождающийся вздрагиваниями,
движениями глазных яблок, сновидениями. В
энцефалограмме появляются волны, напоминающие
реакции внимания при бодрствовании, но более
высокой частоты. Разбуженные в этом состоянии
люди отмечали, что видели сны. Нарушения
парадоксального сна человек переживает тяжело.
Стадии D
и Е
обозначают как
период медленного сна, а стадию Р
– как
период парадоксального сна. В течение ночи
глубина сна может многократно меняться.
Соответственно стадии сна будут сменять друг
друга при выходе из глубокого сна в обратном
порядке, а при следующем его углублении – в
обычной последовательности. Поэтому периоды
медленного и быстрого (парадоксального) сна
много раз чередуются. Обычный ночной сон состоит
из 4–6 завершенных циклов, каждый из которых
начинается с фазы медленного сна и заканчивается
фазой быстрого сна. Длительность цикла
составляет от 60 до 90 мин. При нормальном 8-часовом
ночном сне медленный сон занимает в общей
сложности 6,5 ч, а быстрый – более 1,5 ч.
Стимулами к пробуждению от сна могут
служить: яркий свет, шум, сигналы от внутренних
органов (голодного желудка, переполненного
мочевого пузыря), усиление гормональной
активности и обмена веществ.
Теории сна
По мере накопления факторов и
наблюдений за сном человека и животных возникали
разные теоретические представления о его
природе. Познакомимся с некоторыми их них.
1. Теория гипнотоксинов
. Всем
известное освежающее действие сна наводило на
мысль, что за это время организм освобождается от
накопившейся при дневной деятельности
токсических продуктов обмена веществ, которые и
вызывают сонное торможение нервных клеток мозга.
Недавно было показано участие гуморальных
факторов в развитии сна. Из крови животного,
заснувшего в результате раздражения
определенных зон таламуса, был получен пептид
дельта-сна
, введение которого вызывало сон.
2. Теория центров сна
. Эта
теория ведет начало от клинических наблюдений за
больными энцефалитом, вызывающим летаргический
сон. У этих больных оказывается воспаленным
определенный участок ствола мозга, который стали
рассматривать как центр сна. Предположение, что
сон вызывается возбуждением специальных
центров, получило поддержку в опытах с
раздражением структуры промежуточного мозга,
под действием которого кошка укладывалась в
характерную позу сна и засыпала (рис. 7). Однако
дальнейшие исследования показали, что такой
результат можно получить, раздражая в
определенном режиме стимуляции различные
структуры мозга, что противоречило
представлению о нервном центре, который должен
был иметь определенную локализацию. Кроме того,
клинические наблюдения показали, что патология
сна не связана с определенной локализацией
поражения мозга. Вместе с тем вопрос о центрах
сна представляет существенный интерес.
3. Теория условного торможения
. При
изучении условных рефлексов представителями
школы И.П. Павлова было обнаружено, что
выработка различных видов условного торможения
может приводить ко сну. Это наблюдали при
выработке дифференцировок, запаздывания,
условного тормоза. Аналогичные обстоятельства
вызывают сонливость у человека. Отсюда был
сделан вывод, что «внутреннее торможение
условных рефлексов и сон – один и тот же
процесс».
4. Теория деафферентации сенсорных
систем
. Основанием для этой теории явились
факты развития глубокого сна у животных с
выключенными основными путями поступления
информации в большие полушария (путем
перерезания мозгового ствола на уровне,
предшествующем среднем мозгу). В подкрепление
этой теории приводят описание больного,
сохранившего из всех органов чувств только один
глаз и одно ухо (этот больной засыпал, как только
их закрывали), и эксперименты с хирургическим
выключением у собаки зрения, слуха и обоняния, в
результате чего она почти все время спала.
5. Теории неспецифических
регуляторов сна–бодрствования
. Особую роль в
неспецифической регуляции функционального
состояния высших отделов мозга играет
восходящая активирующая система ретикулярной
формации среднего мозга. Ее раздражение вызывает
реакцию пробуждения, повышает возбудимость коры
мозга. Снижение влияний ретикулярной формации на
кору приводит к развитию сна. Этим объясняется
глубокий непробудный сон после перерезки
мозгового ствола перед средним мозгом.
Продолжение следует
При замыкании и размыкании цепи ток
устанавливается не мгновенно. Эффект
замедления определяется индуктивностью
цепи. Найдём зависимость
при размыкании и замыкании цепи.
При
размыкании цепи ток уменьшается от
значения до нуля и при этом возникает э.д.с.
самоиндукции ,
противодействующая убыванию тока. В
каждый момент времени ток в цепи
определяется законом Ома:
.
Интегрируя уравнение от
до
,
получим:
,
где
- постоянная, имеющая размерность
времени, называется временем релаксации.
Чем больше
,
тем медленнее спадает ток. За времясила тока в цепи уменьшается враз (приблизительно в 3 раза) (см.
зависимость 1 на рисунке).
Если
два электрических контура находятся
вблизи, то они могут влиять друг на
друга. Такие контура называютсяиндуктивно
связанными.
Рассмотрим два таких
контура (см. рисунок). Если по первому
контуру пропустить ток
то магнитный поток, сцепленный со вторым
контуром, будет пропорционален току
,
а также будет зависеть от взаимной
ориентации контуров, их геометрических
размеров, числа витков и магнитных
свойств среды. Можно записать:
.
Здесь коэффициент
называетсявзаимной индуктивностью
второго контура в зависимости от
первого. Обратно, если пропустить токчерез второй контур, то для магнитного
потока сцепленного с первым контуром,
можно записать:
.
Для линейных сред коэффициенты
и равны между собой:
.
Взаимная индуктивность или индуктивность
измеряется в генри (Гн).
Взаимная индуктивность
численно равна магнитному потоку,
сцеплённого с одним из контуров, при
единичном токе в другом контуре. Взаимная
индуктивность зависит от формы, размеров
и взаимной ориентации контуров, магнитной
проницаемости среды.
Например, взаимная индуктивность двух
катушек с общим сердечником равна:
,
где
– объём сердечника,и- число витков на единицу длины образующей
сердечника для первой и второй катушки.
Докажем
это. Пусть на первой катушке течет ток
(см. рисунок). Для достаточно длинной
катушки, пренебрегая краевыми эффектами,
будем считать магнитное поле в сердечнике
однородным:
.
Магнитный поток, сцепленный со второй
катушкой, будет при этом равен:
Откуда и следует выражение для
,
если учесть, что - длина сердечника.
Отметим, что полученное соотношение
для
является приближенным и его можно
представить иначе:
,
где
и- индуктивность катушек.
Если
по одному из контуров пропустить
переменный ток, то во втором возникнет
индукционный ток в соответствии с
законом Фарадея.
Например, если в первом контуре
,
то магнитный поток, сцепленный со вторым
контуром, будет изменяться с течением
времени и в нем возникнет э.д.с. индукции.
.
При
:
.
Очевидно,
.
Возникающее в контурах э.д.с называют
э.д.с. взаимной индукции.
Направление токов и э.д.с. взаимной
индукции определяется правилом Ленца
(см. рисунок)
Возникающий индукционный ток во втором
контуре
своим магнитным полем препятствует
росту магнитного потока от первого
контура.
Возникающий индукционный ток во втором
контуре своим магнитным полем препятствует
уменьшению магнитного потока от первого
контура.
Изменение токов в индуктивно связанных
контурах в линейных средах описывается
законом Ома:
где
- э.д.с. источников в контурах 1 и 2,
- индуктивность контуров, – взаимная индуктивность контуров.
Отметим, что на явлении взаимной индукции
основано действие трансформаторов,
служащих для преобразования токов и
напряжений.
Рассмотрим
холостой ход трансформатора. Это случай,
когда вторичная обмотка трансформатора
не нагружена (см. рисунок). В этом случае
можно записать:
Трансформаторы служат для повышения
или понижения напряжения. Для токов в
обмотках трансформатора наблюдается
обратно пропорциональная зависимость
от числа витков:
.
Обосновать самостоятельно.
Работа 176.
1. Объясните, как работают синапсы.
Когда по аксону нервные импульсы доходятдо синапса, пузырьки с раздражающим веществом лопаются, и жидкость вытекает в синаптическую щель. В зависимости от ее состава клетка возбуждается или тормозится.
2. В чем суть центрального торможения?
Это активный нервный процесс, возникающий в центральной нервной системе и приводящий к угнетению или предупреждению возбуждения.
3. Заполните таблицу.
Работа 177. Заполните таблицу.
Доминанта
Закон взаимной индукции торможения-возбуждения
Мощный очаг временного возбуждения в коре больших полушарий и других отделах мозга, вызванный сильной потребностью, называется доминантной
.
Потребность существует до тех пор, пока не будет удовлетворена или пока другая, более сильная потребность не вытеснит ее.
По закону взаимной индукции доминантный очаг возбуждения затормаживает
все остальные очаги. Возникающее возбуждение переключается на доминантный очаг, который усиливается за его счет. Состояние доминанты облегчает выработку рефлексов
.
возбуждение (или торможение), возникающее в том или ином участке мозга, в конкурирующих центрах вызывает противоположный процесс. Это проявления закона взаимной
индукции
.
В одном и том же центре один и тот же процесс не может существовать вечно, поэтому возбуждение в нем сменяется торможением
, а торможение возбуждением
.
Опъянение вызывает состояние эйфории, которое сменяется тоской и агрессией, т.к. падает уровень глюкозы в крови и происходит эффект "взаимной индукции".
Работа 178. Явление доминанты часто используется в искусстве так же, как и закон взаимной индукции возбуждения и торможения. Привычные образы, идеи, шрифт нередко создают довольно устойчивую доминанту, которая затрудняет формирование конкурирующих образов или идей.
1. Рассмотрите рисунок 107 в учебнике. Почему многие не замечают фигур играющих детей, принимая изображения за извилины больших получшарий головного мозга?
Мы видим то, что привыкли видеть, благодаря сформировавшейся доминанте.
2. Что позволяет нам в трех четрынх пятнах разглядеть фигуру кролика?
Приписка "силуэт кролика".
3. Почему на рисунке трудно заметить фигуру Наполеона?
Потому чт фигура скрыта фоном природы.
4. Почему у лудей, которые четко воспринимают деревья и фигуру Наполеона, эти образы возникают последовательно, сменяя друг друга, или один из них переходит на задний план?
 Р и с. 8. Одновременная индукция: А - положительная; Б - отрицательная
Р и с. 8. Одновременная индукция: А - положительная; Б - отрицательная

 ри
размыкании цепи ток уменьшается от
значения
ри
размыкании цепи ток уменьшается от
значения до нуля и при этом возникает э.д.с.
самоиндукции
до нуля и при этом возникает э.д.с.
самоиндукции ,
противодействующая убыванию тока. В
каждый момент времени ток в цепи
определяется законом Ома:
,
противодействующая убыванию тока. В
каждый момент времени ток в цепи
определяется законом Ома: .
. до
до
 ,
получим:
,
получим:
 ,
, - постоянная, имеющая размерность
времени, называется временем релаксации.
- постоянная, имеющая размерность
времени, называется временем релаксации. ,
тем медленнее спадает ток. За время
,
тем медленнее спадает ток. За время сила тока в цепи уменьшается в
сила тока в цепи уменьшается в раз (приблизительно в 3 раза) (см.
зависимость 1 на рисунке).
раз (приблизительно в 3 раза) (см.
зависимость 1 на рисунке).
 .
. сли
два электрических контура находятся
вблизи, то они могут влиять друг на
друга. Такие контура называютсяиндуктивно
связанными.
Рассмотрим два таких
контура (см. рисунок). Если по первому
контуру пропустить ток
сли
два электрических контура находятся
вблизи, то они могут влиять друг на
друга. Такие контура называютсяиндуктивно
связанными.
Рассмотрим два таких
контура (см. рисунок). Если по первому
контуру пропустить ток
 то магнитный поток, сцепленный со вторым
контуром, будет пропорционален току
то магнитный поток, сцепленный со вторым
контуром, будет пропорционален току
 ,
а также будет зависеть от взаимной
ориентации контуров, их геометрических
размеров, числа витков и магнитных
свойств среды. Можно записать:
,
а также будет зависеть от взаимной
ориентации контуров, их геометрических
размеров, числа витков и магнитных
свойств среды. Можно записать: .
. называетсявзаимной индуктивностью
второго контура в зависимости от
первого. Обратно, если пропустить ток
называетсявзаимной индуктивностью
второго контура в зависимости от
первого. Обратно, если пропустить ток через второй контур, то для магнитного
потока сцепленного с первым контуром,
можно записать:
через второй контур, то для магнитного
потока сцепленного с первым контуром,
можно записать: .
. и
и равны между собой:
равны между собой: .
. численно равна магнитному потоку,
сцеплённого с одним из контуров, при
единичном токе в другом контуре. Взаимная
индуктивность зависит от формы, размеров
и взаимной ориентации контуров, магнитной
проницаемости среды.
численно равна магнитному потоку,
сцеплённого с одним из контуров, при
единичном токе в другом контуре. Взаимная
индуктивность зависит от формы, размеров
и взаимной ориентации контуров, магнитной
проницаемости среды. ,
, – объём сердечника,
– объём сердечника, и
и - число витков на единицу длины образующей
сердечника для первой и второй катушки.
- число витков на единицу длины образующей
сердечника для первой и второй катушки. окажем
это. Пусть на первой катушке течет ток
окажем
это. Пусть на первой катушке течет ток
 (см. рисунок). Для достаточно длинной
катушки, пренебрегая краевыми эффектами,
будем считать магнитное поле в сердечнике
однородным:
(см. рисунок). Для достаточно длинной
катушки, пренебрегая краевыми эффектами,
будем считать магнитное поле в сердечнике
однородным: .
. ,
если учесть, что
,
если учесть, что - длина сердечника.
- длина сердечника. является приближенным и его можно
представить иначе:
является приближенным и его можно
представить иначе: ,
, и
и - индуктивность катушек.
- индуктивность катушек. сли
по одному из контуров пропустить
переменный ток, то во втором возникнет
индукционный ток в соответствии с
законом Фарадея.
сли
по одному из контуров пропустить
переменный ток, то во втором возникнет
индукционный ток в соответствии с
законом Фарадея. ,
то магнитный поток, сцепленный со вторым
контуром, будет изменяться с течением
времени
,
то магнитный поток, сцепленный со вторым
контуром, будет изменяться с течением
времени и в нем возникнет э.д.с. индукции.
и в нем возникнет э.д.с. индукции. .
. :
: .
. .
.
 своим магнитным полем препятствует
росту магнитного потока от первого
контура.
своим магнитным полем препятствует
росту магнитного потока от первого
контура.

 - э.д.с. источников в контурах 1 и 2,
- э.д.с. источников в контурах 1 и 2,
 - индуктивность контуров,
- индуктивность контуров, – взаимная индуктивность контуров.
– взаимная индуктивность контуров. ассмотрим
холостой ход трансформатора. Это случай,
когда вторичная обмотка трансформатора
не нагружена (см. рисунок). В этом случае
можно записать:
ассмотрим
холостой ход трансформатора. Это случай,
когда вторичная обмотка трансформатора
не нагружена (см. рисунок). В этом случае
можно записать: .
. .
. ,
оценим напряжение на вторичной обмотке:
,
оценим напряжение на вторичной обмотке: .
. .
.